
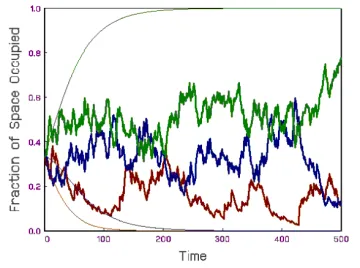
Figure 1: Three hypothetical species competing for space in the lottery model. The thick fluctuating lines demonstrates stable coexistence in a fluctuating environment due the storage effect. Each line is the fraction of space in the system occupied by a particular species as a function of time. The thin smooth lines shows the competitive exclusion that results in the absence of environmental fluctuations.
The storage effect is a general mechanism of species coexistence. It is one of two mechanisms (the other being fitness-density covariance) allowing environmental niche differences to promote diversity. It is involves fundamentally a three way interaction between species, their responses to a varying physical environment, and their responses to competition or to similar multispecies density-dependent processes such as apparent competition. The mechanism is most easily understood in terms of three contributing ingredients:
Species-specific responses to the environment
Responses to the physical environment are changes in population parameters such as the per capita birth rate, per capita death rate, and germination fraction. They can also refer to parameters that are commonly thought of as operating at the individual level, but which are driven by the common environment of a population with consequences at the population level. For example, resource uptake rates, respiration rates, and foraging rates are more often studied the level of the individual but are likely to be correlated over the individuals sharing the same conditions of the physical environment, with important population-level consequences. These responses are species-specific when different species have different patterns of response to the physical environment. One way of measuring these different patterns is through correlation. A large positive correlation indicates low specificity, and a large negative one indicates high specificity. The correlation coefficient, however, does not take into account the magnitude of the variation. The covariance between species would do this, or more usefully, the time by species interaction variance, which is that component of variation in a population parameter that is as specific to a species.
Covariance between environment and competition
Responses to competition are measured as the change in fitness of an individual due to the presence of other individuals of the same or different species with which an individual may compete. Such responses can commonly be measured by removal experiments. Neighborhood competition experiments in plant ecology are one example. Covariance between environment and competition is the statistical covariance between responses to the environment and responses to competition. Due to species-specific responses to the environment, the magnitude and sign of the covariance is expected to vary with the density of a species and the densities of its competitors. In general, a species at high density, dominating a system is likely to have strong positive covariance between environment and competition. These reason is that as the environment becomes increasingly favorable for a species, its demand for resources is likely to increase, and because the species dominates the system, it is likely to have a strong total competitive effect on itself (and also on other species). Its response to competition will therefore covary positively with its response to the environment.
On the other hand, a species at low density will not cause much competition. It experiences competition from other species and thus would be expected to experience high competition when dominant species have favorable environmental conditions. However, due to species-specific responses to the environment, the most favorable environmental conditions for the dominants would not correspond to the most favorable environmental conditions for other species. As a consequence, responses to competition and responses to the environment are not likely to covary as strongly for species at low density as for species at high density. It follows that population growth opportunities occur for low density species when favored by the environment, because this growth is not opposed by competition.
Buffered population growth (aka subadditivity)
Population growth is buffered against unfavorable conditions occurring over time by life-history stages that are long-lived and whose persistence is insensitive, compared to other stages, to varying conditions. The adult individuals of many species tend to be in this category. For species where the adult stage is short-lived–annual and ephemeral species–dormant seeds or eggs often act as buffers. Buffering also occurs in space. When the environment is spatially variable buffering occurs by dispersal between localities in different environmental states.
Buffered population growth is critical to coexistence by the storage effect. Buffering has the effect of skewing growth rate fluctuations in a positive direction because the effects of unfavorable conditions are diminished. This property works in conjunction with the lower covariance between environment and competition experienced by species at low density. Species at low density experience more variable growth conditions, but buffered population growth skews these fluctuations in a positive direction, fostering a species’ recovery from low density. This tendency to favor species with low covariance between environment and competition is the mathematical property of subadditivity. Together with species-specific responses to the environment, and covariance between environment and competition, it forms the storage-effect coexistence mechanism.
References
Chesson, P.L., Warner, R.R. 1981. Environmental variability promotes coexistence in lottery competitive systems. American Naturalist 117, 923-943.
Chesson, P.L. 1985. Coexistence of competitors in spatially and temporally varying environments: a look at the combined effects of different sorts of variability. Theor. Pop. Biol. 28, 263-287.
Chesson, P., Huntly, N. 1989. Short-term instabilities and long-term community dynamics. Trends in Ecology and Evolution 4, 293-298.
Chesson, P. 1990. Geometry, heterogeneity and competition in variable environments. Phil. Trans. Roy. Soc. Lond, Series B 330, 165-173.
Hatfield, J., Chesson, P. 1996. Multispecies lottery competition: a diffusion analysis. Pp 615-622 in S. Tuljapurkar and H. Caswell, eds. Structured Population Models in Marine, Freshwater, and Terrestrial Systems. Chapman and Hall, New York.
Chesson, P., Pacala, S., Neuhauser, C. 2001. Environmental niches and ecosystem functioning. Pages 213-245 in "The Functional Consequences of Biodiversity", Ann Kinzig, Stephen Pacala, and David Tilman, eds, Princeton University Press.
Gebauer, R. L. E., Schwinning, S., Huntly, N., Wiegand, K., Ernest, M. S. K., Sher, A., Novoplansky, A., and Weltzin, J.F. 2004. Resource pulses, species interactions and diversity maintenance in arid and semi-arid environments. Oecologia